
[Cell Biology] Lịch sử phát triển học thuyết màng tế bào
Vào 3,7 tỉ năm trước, ở dưới lòng đại dương trong những hố nitơ nguyên thủy, sự kết hợp giữa những nguyên tố hóa học bằng một sự ngẫu...

Vào 3,7 tỉ năm trước, ở dưới lòng đại dương trong những hố nitơ nguyên thủy, sự kết hợp giữa những nguyên tố hóa học bằng một sự ngẫu nhiên nào đó đã hình sự sống đầu tiên trên trái đất. Khi đó, sự sống bắt đầu từ cấu trúc giản đơn là RNA đơn lẻ trôi nổi trong lòng đại dượng và chúng có thể tự sao và nhân đôi tạo ra cá thể mới. Nhưng để tạo ra một cấu trúc phức tạp và đa dạng như trái đất hiện nay, chúng cần phải được lưu trữ cũng như tránh những tác nhân phá hủy bên ngoài từ đó màng tế bào được hình thành. Tuy rằng, màng tế bào được hình thành rất sớm từ thuở sơ khai hình thành trái đất, phải đến rất lâu sau những cuộc cách mạng khoa học chúng ta mới phát hiện ra tế bào qua lớp kính hiển vi từ đó tạo tiền đề cho những khám phá và hiểu biết về những thế giới siêu tí hon này.
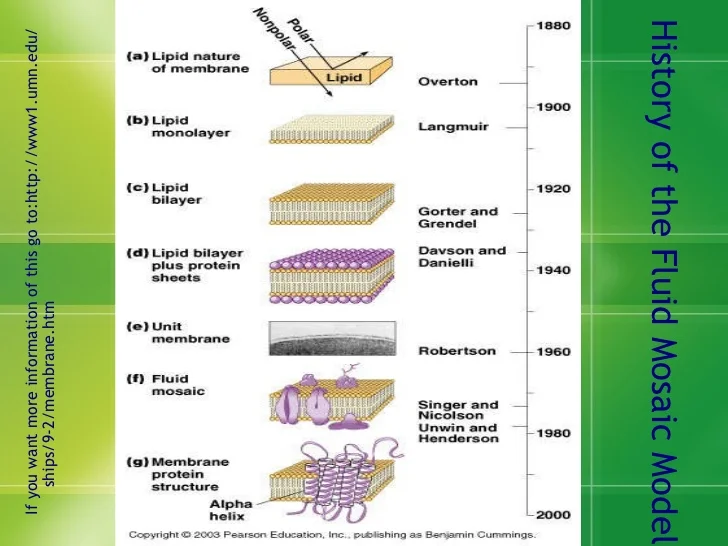
Đây là biểu đồ biểu thị những phát hiện và phát về màng tế bào theo thời gian (mình sẽ đi vào từng thời kỳ của phát triển về học thuyết màng tế bào).
I) Overton và Langmuir: Lipids là một hợp chất quan trọng của màng tế bào

Một điểm khởi đầu thuận lợi cho khám phá của chúng ta là những nguyên cứu đầu tiên của nhà tiến sĩ người nga Charles Ernest Overton trong những năm thập niên 90 của thế kỷ 19. Khi làm việc với rễ những gốc cây, ông đã quan sát thấy rằng các vật chất lipid hòa tan có thể thâm nhập và tế bào trong khi các vật chất nước hòa tan thì không. Từ quan sát trên, Overton đã kết luận rằng lipid hiện diện trên bề mặt của tế bào như một chiếc ‘áo khoác’. Ông còn cho rằng chiến áo khoác này của tế bào có thể là một hỗn hợp cấu thành từ cholesterol và lecithin, tầm nhìn sáng suốt này của ông đã hỗ trợ cho chúng ta đã hiểu thêm về sự quan trọng của sterol và phophoslipid-những hợp chất cấu thành nên màng tế bào.
Bước cải tiến thứ hai được phát hiện khoảng một thập kỷ sau qua công trình nguyên cứu của Irving Langmuir, ông đã quan sát qua việc thanh lọc phospholipid bằng cách phân hủy chúng trong benzene và phân lớp mẫu benzene-lipid trên mặt nước. Khi benzene bay hơi, những phân tử còn lại là một lớp lipid đặc, còn gọi là ‘monolayer’ (lớp đơn). Và bởi vì phospholipid là chất lưỡng tính nên Langmuir đã giải thích rằng phospholipid sẽ định hướng cho những vật chất ưa nước (hydrophilic) nằm ở bền mặt tế bào còn đuôi sợ nước (hydrophobic) sẽ tránh xa khỏi bề mặt. Phát hiện về lớp lipid đơn tầng của Langmuir đã trở thành một trong tiền đề cơ bản cho những bước tiến xa hơn về hiểu biết của chúng ta về màng tế bào ở thế kỷ 20.
II) Gorter và Grendel: Cấu trúc cơ bản của màng tế bào là một lớp lipid đôi.

Bước tiến lớp tiếp theo là vào những năm 1925, khi hai nhà vật lý học Hà Lan, Evert Gorter và F.Grendel, đã tách chiếc lipid từ một lượng erythrocytes (tế bào hồng cầu) và sử dụng phương pháp Langmuir để phân lipid như một lớp đơn trên mặt nước. Và kết quả cho họ thấy rằng đây không phải là một mà hai lớp lipid so với tổng bề mặt của tế bào. Theo đó họ kết luận rằng màng tế bào hồng cầu bao gồm hai lớp lipid.
Về giả thuyết về cấu trúc lớp đôi, Gorter và Grendel đã giải thích rằng sẽ thuận lợi hơn về mặt nhiệt động lực hơn nếu chuỗi không phân cực của hydrocarbon của từng lớp hướng vào trong, cách xa khỏi môi trường nước ở bên trong bề mặt của tế bào và nhóm phân cực ưa nước sẽ nằm ở môi trường ngoài. Thí nghiệm và kết luận của Gorter và Grendel đã mở ra cách cửa đầu tiên cho nỗ lực các nhà khoa học cho sự hiểu biết màng tế bào ở góc độ phân tử.
III) Davson và Danielli: Màng tế bào bao gồm cả proteins.

Sau khi Gorter và Grendel đăng mô hình của họ về lớp lipid đôi, sự việc trở nên rõ ràng khi lớp lipid đôi không thể giải đáp được cho toàn bộ tính chất của màng tế bào- cụ thể là sức căng bề mặt, tính thấm-tan và kháng điện. Ví dụ như, sức căng bề mặt của lớp lipid có thể bị điều chỉnh nhờ việc thêm vào các protein. Hơn nữa, khi đường, ion và các vật chất hòa tan với nước di chuyển vào trong tế bào xuyên qua lớp lipid đôi vốn là không thể nào vì tính chất của chúng.
Để giải thích cho câu hỏi này, Hugh Davson và James Danielli cho rằng có sự xuất hiện của proteins trên màng. Trong năm 1935, họ đã đề xuất rằng màng tế bào về mặt sinh học bao gồm lớp lipid đôi được bao phủ bởi một tấm protein mỏng. Mô hình của họ như một chiếc bánh ’sandwich’ với protein-lipid-protein, là mô tả chi tiết đầu tiên về sự hiện diện của hệ thống màng và đã thống trị suy nghĩ của các nhà sinh học tế bào trong những thập kỷ sau.
Tuy vậy bản thiết kế vẫn được chỉnh sửa để phù hợp cho những phát hiện mới. Một trong những đề xuất đáng chú ý là vào năm 1954, các protein ưa nước có thể thâm nhập và màng tế bào để hỗ trợ việc hình thành các ‘hố phân cực’ thông qua lớp lipid đôi kị nước. Những protein có thể cho phép những vật chất hòa tan trong nước đi xuyên qua màng tế bào. Đặc biệc là lớp lipid bên trong màng chiếm hầu như toàn bộ vật chất kị nước của màng và những hợp chất protein đã giải thích cho tính chất ưa nước của chúng.
Điều quan trọng và đáng chú ý nhất trong mô hình của Davson và Danielli, nó đã nhấn mạnh tầm quan trọng của protein ở cấu trúc màng và là tiến đề cơ bản cho những nguyên cứu về cấu trúc màng sau này.
IV) Roberton: Tất cả màng tế bào đều chia sẻ một cấu trúc chung cơ bản.

Với sự ra đời của kính hiển vi điện tử vào những năm thập niên 50 của thế kỷ 20, các nhà sinh học tế bào có thể kiểm chứng sự hiện diện của màng xung quanh tế bào. Họ cũng quan sát được rằng sự hiện diện những bào quan cũng sẽ bị ràng buộc bởi những loại màng cơ bản. Thêm vào đó, khi thử nghiệm cho màng khi bị được đánh dấu bởi osmium, một kim loại nặng, rồi được phóng đại lên, họ đã nhìn thấy một vùng rộng lớn của cấu trúc ‘đường sắt’ xuất hiện như hai đường kẽ đen phân tách bởi một vùng trung gian, với độ dày khoảng 6-8 nm (hình ở trên). Khi nhìn ta có thể thấy rằng hai màng tế bào được phân tách ra bởi một vùng gian bào mỏng. Bởi vì mẫu này xuất hiện phổ biến ở nhiều loại tế bào khác nhau, J. David Robertson đã cho rằng tất cả các loại màng tế bào đều chia sẻ một cấu trúc chung được ông ấy gọi là ‘unit membrane’ (đơn vị màng)
Khi được công bố, đơn vị cấu trúc màng được công nhận rộng rãi với mô hình Davson-Danielli. Robertson còn cho rằng vùng gian bào (nằm giữa hai đường kẻ sọc đen của mô hình tam lớp) chứa vùng sợ nước của phân tử lipid, vùng không màu. Ngược lại, hai đường kẽ đen được cho là nhóm đầu phospholipid và tấm protein gắn với bề mặt, bởi vì sự tương thích của chúng với kim loại nặng. Cách diễn giải này hỗ trợ mạnh mẽ cho quan sát của Davson-Danielli rằng màng tế bào gồm một lớp lipid đôi được khoác lên bởi một tấm protein mỏng ở hai bề mặt.
V) Những phát hiện mới để lộ ra sự thiếu hụt trong mô hình Davson-Danielli
Cho dù mô hình nhận được sự chấp thuận qua quan sát kính hiển vi và từ nghiên cứu mở rộng của màng bởi Robertson, mô hình Davson-Danielli gặp những khó khăn vào những năm 1960s khi ngày càng nhiều dữ liệu xuất hiện không đồng nhất với mô hình của họ. Dựa theo kính hiển vi, hầu hết màng dày từ 6-8nm và lớp lipid đôi dày 4-5nm. Thế là chỉ còn 1-2nm trên bề mặt của màng đủ chỗ cho proteín, một khoảng trống vừa đủ cho một lớp mỏng của protein. Nhưng sau khi proteins được để riêng và nghiên cứu, nó hầu như rõ ràng là Globular proteins (protein dạng cầu) với kích thước và hình dạng không nhất quán với ý tưởng về một lớp proteins mỏng trên hai bề mặt của màng.
Thêm vào vấn đề phức tạp này, mô hình Davson-Danielli không nghĩ đến sự riêng biệt của từng loại màng. Tùy theo nguồn gốc, màng sẽ đa dạng theo cấu trúc hóa học và đặc biệt với tỉ lệ protein-lipid, có thể là ba hoặc lớn hơn trong vài tế bào vi khuẩn, hoặc là bao meylin bao bọc dây thần kinh có thê có tỉ lệ nhỏ đến 0.23. Vì thế hai màng từ hai ti thể sẽ có sự khác nhau: tỉ lệ protein-lipid tầm 1.2 cho màng ngoài và tầm 3.5 cho màng trong, nơi chứa toàn bộ enzymes và protein liên quan đến vận chuyển electron và tạo ATP. Nhưng toàn bộ màng đều nhìn giống nhau khi được quan sát dưới kính hiển vi.
Mô hình Davson-Danielli còn được xem xét lại qua việc nghiên cứu màng khi được tác dụng với phospholipases, là enzymes phân hủy Phospholipids bằng cách gỡ bỏ phẩn đầu. Dựa theo mô hình nhóm đầu ưa nước của màng tế bào nên được phủ bởi một lớp protein và vì thế bảo vệ bởi sự phân hủy của phospholipase. Nhưng, đến 75% màng phospholipid có thể bị phân hủy khi bị tác động bởi phospholipases, từ đó dẫn đến kết luận hầu hêt phần đầu của phospholipid đều ở trên bề mặt của màng và không được bảo vệ bởi lớp protein.
Hơn nữa, mô hình Davson-Danielli không nhận được sự hỗ trợ từ các nhà khoa học những người cố gắng cô lập những protein trên vì mô hình của ông không đề cặp đến vị trí của protein trên bề mặt màng. Hầu hết protein của màng được phát hiện là không tan được trong nước và cách duy nhất để tách nó ra là bằng cách dùng dung môi hoặc chất tẩy rửa. Từ những quan sát trên cho thấy rất nhiều protein của màng là chất kị nước (hoặc là chất lưỡng phần) và cho thấy rằng những protein đó hoặc một phần của nó ở trong vùng kị nước của màng nhiều hơn là trên bề mặt.
VI) Singer and Niolson: Một màng chứa dạng khảm lỏng của lipid lớp đôi.

Những vấn đề trước đây với mô hình Davson-Danielli đã kích thích một lượng lớn sự quan tâm trong việc phát triển những ý tưởng mới về tổ chức màng, trong đó đỉnh điểm là vào 1972 với mô hình màng chứa dạng khảm lỏng được đề xuất bởi S.Johnathan Singer và Garth Nicolson. Mô hình này, được biết là đã và đang thống trị quan điểm về cách chúng ta nhìn nhận tổ chức màng, gồm hai đặc điểm chính, đều gắn liền với tên nó. Nói một cách đơn giản, mô hình hình dung một màng như một mảng ghép của các protein được khảm vào, hoặc ít nhất là được gắn vào một cấu trúc dạng lỏng lipid lớp đôi của mô hình cũ nhưng lại nhìn nhận màng protein theo một cách hoàn toàn khác – không phải như những tấm mỏng trên bề mặt màng mà là những thực thể hình cầu rời rạc bên trong lớp lipid kép.
Cách nghĩ về protein màng đã mang tính cách mạng khi Singer Nicolson lần đầu tiền đề xuất nó, nhưng hóa ra nó lại khá phù hợp với dữ liệu. Ba “họ” protein màng được phân biệt dựa trên sự khác biệt trong cách protein được liên kết với lớp đôi. Các màng bào tương được nhúng trong lớp lipid, nơi chúng được giữ cố định bởi lực liên kết hấp dẫn của các phân đoạn sợ nước đối với phần bên trong kị nước của lớp lipid. Các protein ngoại vi ưa nước hơn nhiều và do đó nằm trên bề mặt của màng, nơi chúng được liên kết không hóa trị với các nhóm đầu phân cực của phospholipid và/hoặc với các phần ưa nước của các protein màng khác. Các protein giữ lipid về cơ bản là các protein ưa nước và do đó chúng nằm trên bề mặt màng, nhưng lại được gắn cộng hóa trị với các phân tử lipid được nhúng trong lớp đôi.
Chất lỏng tự nhiên của màng là điểm quan trọng thứ hai của mô hình Singer-Nicolson. Thay vì khóa chặt tại chỗ, hầu hết các thành phần lipid của màng đều chuyển động liên tục, có khả năng di chuyển theo chiều (tức là chuyển động song song với bề mặt màng). Nhiều protein màng cũng có thể di chuyển theo chiều bên trong màng, mặc dù số protein được gắn vào các yếu tố cấu trúc như bộ xương tế bào ở bên này hoặc bên kia của màng và do đó hạn chế khả năng di chuyển của chúng.
Điểm mạnh chính của mô hình khảm chất lỏng là dễ dàng giải thích hầu hết những lời chỉ trích về mô hình Davson-Danielli. Ví dụ, khái niệm về protein được nhúng một phần trong lớp lipid kép phù hợp với bản chất kị nước và cấu trúc hình cầu của hầu hết các protein màng và loại bỏ sự cần thiết phải chứa các protein màng trong các lớp bề mặt mỏng có độ dày không thay đổi. Hơn nữa, sự khác nhau về tỷ lệ protein/lipid giữa các màng khác nhau đơn giản có nghĩa là các màng khác nhau khác biệc về số lượng protein mà chúng chứa. Ngoài ra, sự tiếp xúc của các nhóm đầu lipid ở bề mặt màng rõ ràng là tương thích với tính nhạy cảm của chúng với quá trình xúc tác tiêu hóa của phospholipase, trong khi tính lưu động của các lớp lipid và sự xen kẽ của lipid và protein trong màng giúp dễ dàng hình dung tính di động của chúng.
VII) Unwin và Henderson: Hầu hết các protein màng chứa các đoạn xuyên màng.

Hình minh họa tiếp theo trong dòng thời gian mô tả một tính chất quan trọng của protein màng bào tương mà các nhà sinh học tế bào bắt đầu tìm hiểu vào những năm 1970: hầu hết các protein như vậy có cấu trúc cơ bản là một hoặc nhiều chuỗi kị nước kéo dài lớp lipid kép. Ví dụ trong hình trên là bacteriorhodopsin, protein màng đầu tiên được chứng minh là có đặc điểm cấu trúc này. Bacteriorhodopsin là một protein màng sinh chất được tìm thấy trong vi khuẩn cổ thuộc giống Halobacterium, nơi mà sự hiện diện của nó cho phép tế bào thu năng lượng trực tiếp từ ánh sáng mặt trời. Nigel Unwin và Richard Henderson đã sử dụng kính hiển vi điện tử để xác định cấu trúc ba chiều của bacteriorhodopsin bao gồm một chuỗi peptit đơn lẻ được gấp đi gấp lại qua lớp lipid kép tổng cộng bảy lần. Mỗi đoạn trong số bảy đoạn xuyên màng của protein là một chuỗi xoắn alpha gói chặt chẽ lại bao gồm chủ yếu là các axit amin kị nước. Các đoạn xuyên màng kế tiếp nhau được liên kết với nhau bằng các vòng ngắn của các axit amin ưa nước kéo dài vào hoặc nhô ra khỏi bề mặt phân cực của màng. Dựa trên các nghiên cứu tiếp theo trong nhiều phòng thí nghiệm, các nhà sinh học màng hiện nay tin rằng tất cả các protein xuyên màng đều được gắn trong lớp lipid kép bởi một hoặc nhiều đoạn xuyên màng.
VIII) Những phát hiên gần đây cũng cố thêm về sự hiểu biết của chúng ta về cấu trúc của màng tế bào
Từ khoảnh khắc Singer và Nicoloson đề xuất Mô hình khảm chất lỏng đã tạo ra cuộc cách mạng làm thay đổi cách nhà khoa học nghĩ về cấu trúc màng tế bào. Mô hình đã tạo ra kỉ nguyên mới trong trong việc nghiên cứu màng tế bào, mô hình không chỉ chứng minh được cho mô hình cơ bản mà còn cải tiến và mở mộng thêm. Hơn nữa, sự hiểu biết của chúng ta về cấu trúc của màng được tiếp tục mở rộng cùng với những phát kiến mới củng cố thêm mô hình cơ bản.
Với sự phát triển gần đây nhấn mạnh về ý tưởng màng không phải là cấu trúc tự do, đồng nhất. Cả lipids và proteins đều được nhận lệnh từ màng tế bào và sự điều khiển thường xuyên xảy ra trong màng tế bào, và sự sắp xếp này thường xảy ra ở miền vi động biết biết là lipid rafts, cụ thể hơn về phần này sẽ được đề cập sau những bài viết khác của mình. Thực tế, hầu hết các quá trình tế bào có liên quan đến màng tế bào phụ thuộc rất nhiều về cấu trúc nhất định và phức tạp của lipids và proteins trong màng. Sự liên kết giữa proteins màng và một lipid nào đó có thể là phản ứng riêng biệt và thường quan trọng cho cấu trúc và chức năng của protein trong màng.
Vậy để hiểu hơn về quá trình liên kết màng, chúng ta cần nhiều hơn một mô hình khảm lỏng truyền thống với lipids và proteins trôi nổi tự do xung quanh. Nhưng mô hình này vẫn là hiểu biết cơ bản của chúng ta về cấu trúc màng, nên vẫn cần thiết để xem xét về những đặc trưng của nó. Những đặc trưng này bao gồm hóa học, sự phân bổ không đối xứng, và tính lưu động của màng lipid; mối quan hệ giữa proteins với màng lipid đôi; và tính linh động của chúng trong màng.
Chú thích thêm:
Chất ưa nước: Chất ưa nước là một phân tử hoặc thực thể phân tử khác bị thu hút bởi các phân tử nước và có xu hướng tan trong nước.
Đuôi kị nước: Chất kị nước không bị hấp dẫn bởi nước và có thể bị đẩy bởi nước.
Vùng gian bào: Là khu vực đệm giữa lòng mạch và tế bào, nhờ vậy những biến động lớn từ lòng mạch không ảnh hưởng trực tiếp ngay đến khu vực tế bào, màng tế bào ngăn cách hai khu vực này.
Hố phân cực: Sau này còn được gọi là kênh protein, dùng để vẫn chuyển các chất ưa nước đi qua màng tế bào
Khảm lỏng: là bề mặt có tính lỏng và được khảm lên.
Đây là một bài dịch của mình trong series sinh học tế bào, vì mình dịch hoàn toàn theo ngẫu hứng nên các bài nhiều lúc sẽ khá riêng rẽ và ít liên kết. Nhưng nếu khi nhìn với góc độ từ vi mô đến vĩ mô thì tất cả mọi thứ về tế bào sẽ đều có mối liên kết và quan hệ đến nhau. Mong các bạn thông cảm.
Mình không biết khi nào mình sẽ viết bài tiếp theo, nhưng mong mình sẽ có động lực và năng lượng để tiếp tục làm về series này. Các bạn ủng hộ cho mình một upvote nhé vì nó giúp mình rất nhiều! ^^
Và cũng cảm ơn ba bạn Lê Hoàng, Văn Huy và Hoàng Anh đã giúp đỡ mình rất nhiều để hoàn thiện bài
Link ảnh:
Nguồn của bài mình lấy từ sách: Becker's World of the Cell 8th
Đây là link PDF mình tải sách về: https://fdocuments.in/document/beckers-world-of-the-cell-8th-edition-pdf-beckers-world-of-the-cell-8th-edition.html

The Bard
@jannastarguardian
Science2vn
/science2vn
Bài viết nổi bật khác
- Hot nhất
- Mới nhất